




Developmental Neurobiology Unit
Professor Ichiro Masai

Abstract
Developmental neurobiology unit uses zebrafish as an animal model and will elucidate genetic program that regulate eye development. Specific research projects are to elucidate:
(1) Mechanisms that regulate retinal cell differentiation and neural circuit formation
(2) Mechanisms that regulate photoreceptor degeneration and regeneration
(3) Mechanisms that regulate lens fiber differentiation
(4) Mechanisms that regulate zebrafish behaviors linked to fear/anxiety response
During vertebrate development, the retina is originally derived from anterior neural plate. In this region, six major classes of retinal neurons differentiate and form neural circuits responsible for vision. Thus, the retina provides a good model for studying cell differentiation and neural circuit formation in the developing brain.
First, we will investigate mechanisms that regulate cell differentiation and neural circuit formation.
Second, we will investigate mechanisms that regulate photoreceptor degeneration and regeneration. Photoreceptor degeneration is an important topic for medical research, because more than 300 genetic mutations are associated with photoreceptor degeneration in humans. We focus on zebrafish mutants, in which photoreceptor degeneration is caused by defects in protein synthesis in ER, protein transport from ER to the apical photoreceptive membrane region called the outer segment, and phototransduction. Through these mutant analyses, we will determine how photoreceptors monitor abnormalities in cellular functions and trigger apoptosis. When the zebrafish retina suffers from photoreceptor degeneration, one type of retinal glial cells, namely Müller cells, start to proliferate and generate retinal neurons as neural stem cells. Recent studies revealed that brain-resident immune cells, namely microglia, play an important role in this regeneration program of Müller cells. We will investigate the role of brain-resident immune cells, namely microglia in photoreceptor degeneration and regeneration.
Third, we will investigate lens development, especially, mechanisms that regulate lens fiber differentiation. Through these projects, we will establish key concepts that govern development of multicellular organisms and also contribute to our understanding of pathological processes of human retinal diseases.
In addition to research project on ocular development above, we have collaborated with Greg Stephens unit to investigate behavioral mechanisms that regulate social interaction such as fish shoaling and fear/anxiety response using zebrafish as a model.
1. Staff
- Dr. Yuko Nishiwaki, Group leader
- Dr. Yuki Takeuchi, Staff scientist
- Dr. Wei-Chieh Chiang, Staff scientist
- Dr. Luis Carretio, Staff scientist
- Dr. Mai Omar Abdulrahman Ahmad, Junior Research Fellow (JRF) (–26 July 2022)
- Dr. Swathy Babu, Junior Research Fellow (JRF) (–25 May 2022)
- Dr. Manana Kutsia, phD student and Junior Research Fellow (JRF) (–28 December 2022)
- Mr. Bedish Chatterjee, phD student
- Mr. Hung-Ju Chiang, phD student
- Mr. Dongpeng Hu, phD student
- Ms. Darshini Ravishanker, phD student
- Mr. Akira Kawano, phD student
- Ms. Mariia Pavelchenko, phD student
- Mr. Tatsuo Izawa, Rotation student (September 1 -December 28, 2022)
- Mr. Taimu Masaki, Research Intern (November 5, 2021 – May 31, 2022)
- Ms. Jana De Troyer, Research Intern (October 6, 2022 – February 7, 2023)
- Ms. Lakshmi Sowmya Emani, Research Intern (October 6, 2022 – December 20, 2022)
- Mr. Yutaka Kojima, Technician (–30 September 2022)
- Dr. Tetsuya Harakuni, Technician
- Dr. Mamoru Fujiwara, Technician
- Mr. Taichi Kawakami Technician (tempo staff) (April 18, 2022–)
- Ms. Paula Niejenhuis Technician (February 1, 2023–)
- Ms. Moe Inafuku, Laboratory Assistant
- Ms. Chitose Mizuta, Laboratory Assistant
- Ms. Rui Inoue, Laboratory Assistant
- Ms. Xiujun Huang, Laboratory Assistant (–31 December 2022)
- Ms. Yuzuki Ikema, Laboratory Assistant
- Ms. Madoka Makiya, Research Assistant (Tempo staff)
- Ms. Miki Kitamura, Research Assistant (Tempo staff –> OIST staff from February 1, 2023)
- Ms. Kaori Sugiura, Research Assistant (Tempo staff) (4 January 2023–)
- Ms. Mizuki Otake, Research Unit Administrator/Secretary
2. Collaborations
2.1 The Title or Name or Topic of the Collaboration
- Theme: In vivo functional analysis of hypoxia response genes using the zebrafish retina
- Type of collaboration: Joint research agreement
- Researchers: Dr. Ichiro Masai (Developmental neurobiology unit, OIST), Dr. Masayuki Matsushita (Department of Medicine, Ryukyu University)
2.2 The Title or Name or Topic of the Collaboration
- Theme: Role of zebrafish BANP protein in tumor suppression of melanoma
- Type of collaboration: Joint research agreement
- Researchers: Dr. Ichiro Masai (Developmental neurobiology unit, OIST), Dr. Yutaka Kikuchi (Department of Biological Science, Hiroshima University)
3. Activities and Findings
3.1 Mechanism of neural circuit formation in zebrafish retinas
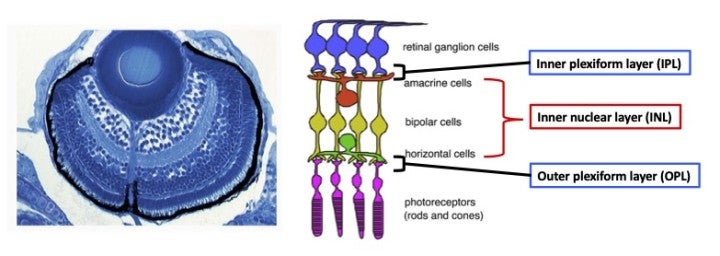
During development, the retina is originally derived from the anterior neural plate. In the retinal region, six major classes of neurons and one types of glial cells differentiate to form the neural circuit, which is responsible for phototransduction and visual processing (Fig. 1). The retinal neural circuit processes visual signals through two synaptic neuropils. At the apical side, the outer plexiform layer (OPL) harbors synapses that transmit input from photoreceptors (PRs) in the outer nuclear layer (ONL) to bipolar (BP) and horizontal cells (HCs) in the inner nuclear layer (INL). At the basal side of the retina, the inner plexiform layer (IPL) is densely packed with synaptic connections formed between BPs and amacrine cells (ACs) in the INL, and retinal ganglion cells (RGCs) in the ganglion cell layer (GCL). The retina contains one type of glial cells called Müller glia (MGs), which span the apico-basal axis of the retina. However, molecular mechanism underlying retinal cells differentiation and neural circuit formation remains to be elucidated. In FY2022, we focused on two zebrafish mutants carrying a genetic mutation in strip1 and banp gene, for studying the mechanism of IPL formation and cell-cycle regulation, respectively.
3.1.1 The role of Strip1 in inner synaptic layer formation
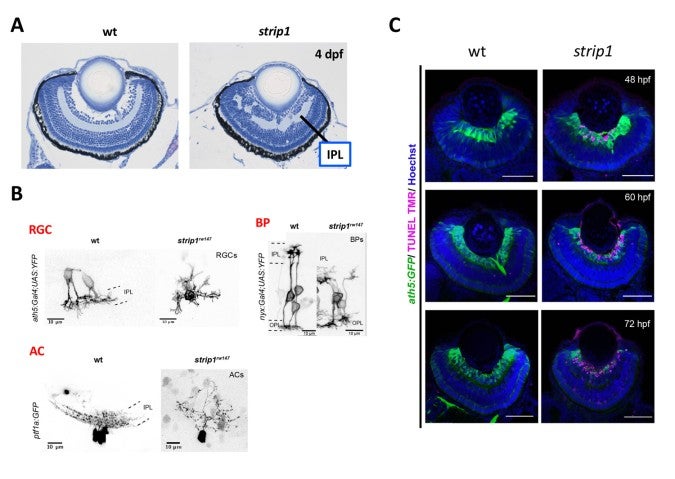
Zebrafish strip1 mutants were identified by zebrafish large-scale mutant screening, which we conducted from 1998 to 2004 (Masai et al., 2003). strip1 mutants shows defects in retinal layer organization, especially IPL (Fig. 2A). IPL is formed by three retinal neurons, RGCs, ACs and BPs. These neurons show an abnormal patterning of dendrites and axons, suggesting that Strip1 is required for proper neurite patterning of these neurons (Fig. 2B). Our cell transplantation analysis revealed that neurite patterning of ACs and BPs is regulated in a cell-non-autonomous manner, indicating that Strip1 does not function in ACs and BPs but rather in RGCs. Indeed, RGCs undergo apoptosis during their maturation in strip1 mutants, which allows ACs and BPs to infiltrate into dying RGC layer (Fig. 2C). This abnormal infiltration of ACs and BPs into RGC layer causes defects in IPL formation. Thus, Strip1 is primarily required for RGC survival.
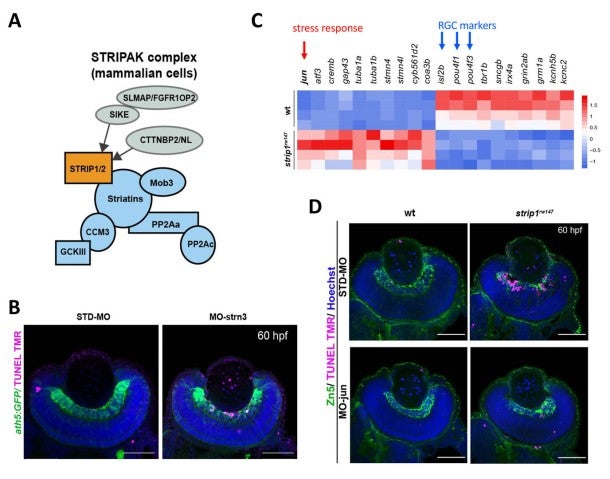
Strip1 was initially identified as a Striatin interacting protein. Recent studies revealed that Strip1 and Striatin cooperate to form a large protein complex called STRIPAK complex (Fig. 3A). We confirmed that zebrafish Strip1 physically interacts with other component of STRIPAK complex including Striatin3 and that knockdown of Striatin3 shows a similar RGC apoptosis. So, Strip1 and Striatin3 function in RGC survival (Fig. 3B). Our bulk-RNA-seq analysis revealed that the JNK/Jun pathway is abnormally activated in RGCs of strip1 mutants (Fig. 3C). Knockdown of Jun rescues RGC apoptosis in strip1 mutants (Fig. 3D). Thus, STRIPAK complex promotes RGC survival by suppressing JNK/Jun-mediated apoptotic pathway. Since RGC death is associated with optic nerve damage, for example, human glaucoma, we compared RNA expression profile between zebrafish strip1 mutants and zebrafish adult glaucoma model with optic nerve injury. Interestingly, Jun activation is also observed in zebrafish glaucoma model, suggesting that the mechanism underlying Strip1-mediated RGC apoptosis provides an important insight on therapeutic development of human glaucoma.
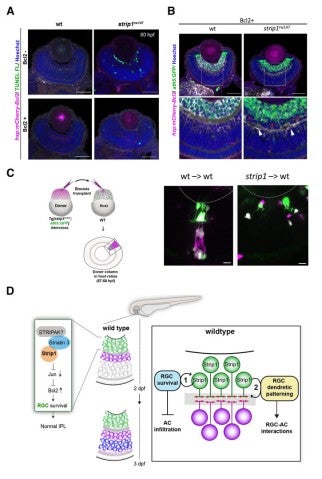
Lastly, to elucidate whether IPL formation defects in strip1 mutants depend on only RGC death, we prevented RGC apoptosis in strip1 mutants by overexpression of anti-apoptotic protein, Bcl2. Bcl overexpression rescued RGC death in strip1 mutants (Fig. 4A). We found that AC infiltration into RGC layer is inhibited in Bcl2-overexpressing strip1mutants; however, IPL fails to be formed at the interface between RGCs and ACs but formed at the intermediate position within AC layer (Fig. 4B). We also confirmed that these survived strip1 mutant RGCs show an abnormal patterning of dendrites and axons (Fig. 4C), indicating that Strip1 is required for proper neurite patterning of RGCs independent of RGC survival, which allows RGCs interacts with ACs to initiate IPL formation program. These data suggest dual roles of Strip1 in IPL formation: (1) RGC survival and (2) RGC neurite patterning (Fig. 4D).
3.1.2 The role of Banp in cell-cycle regulation in the developing retina
Btg3-associated nuclear protein (Banp) was originally identified as a nuclear matrix-associated region (MAR)-binding protein and it functions as a tumor suppressor. At the molecular level, Banp regulates transcription of metabolic genes via a CGCG-containing motif called the Banp motif. However, its physiological roles in embryonic development are unknown. Zebrafish banp mutants were identified by large-scale zebrafish mutant screening, which we conducted from 1998 to 2004 (Masai et al., 2003) (Fig. 5A). We found that banp mutants show mitotic cell accumulation and apoptotic cell death in the developing retina (Fig. 5B). Next, bulk-RNA-seq analysis revealed that p53 dependent-DNA damage response regulators are significantly upregulated in banp mutants (Fig. 5C). Indeed, retinal apoptosis in banp mutants was significantly rescued by p53 knockdown. Furthermore, expression of g-H2AX, which is a global marker of stalled replication forks and DNA double stranded break, was significantly elevated in banp mutant retina (Fig. 5D). These data suggest that DNA replication stress and subsequent DNA damage occur in the absence of Banp, which subsequently activates p53-mediated DNA damage response, leading to upregulation of a cell-cycle arrest gene, p21, and pro-apoptotic genes such as Puma.
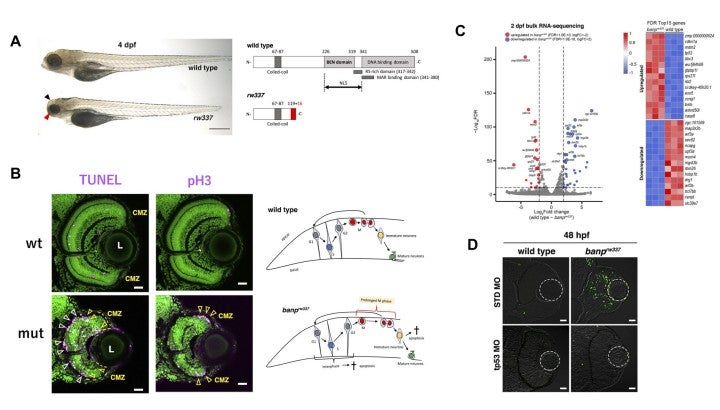
However, a substantial fraction of accumulated mitotic cells in banp mutants is independent of p53. RNA-seq analysis revealed that two chromosomal segregation regulators, Cenpt and Ncapg, are downregulated in banp mutants (Fig. 5C). Indeed, quantitative RT-PCR confirmed their downregulation in banp mutants (Fig. 6A). ATAC-seq revealed that chromatin accessibility is decreased near the transcription start site of both genes in banp mutants (Fig. 6B). It was reported that knockdown of Cenpt and Ncapg causes defects in chromosomal segregation during mitosis in zebrafish (Hung et al., 2017; Seipild et al., 2009). Consistently, mitosis did not proceed smoothly in banp knockdown retinas: it took longer from chromosomal condensation in prophase to chromosomal alignment at the division plane in anaphase (Fig. 6C). Furthermore, aligned chromosomes were often dispersed during chromosomal segregation by the mitotic spindle (Fig. 6D). So, the percentage of cells with abnormal mitosis is significantly higher in banp mutant retinas than those of wild-type siblings. Thus, Banp regulates chromosomal segregation during mitosis.
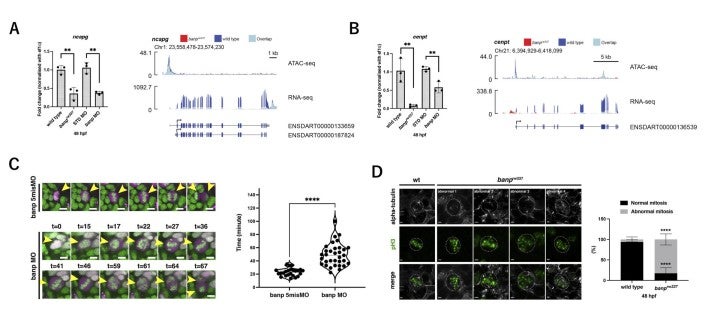
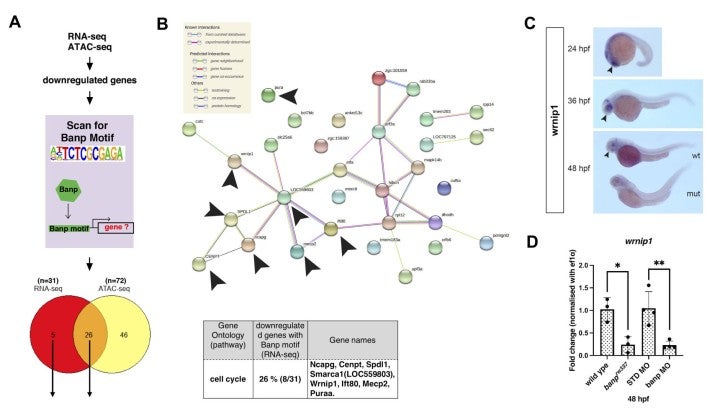
Recently, it was reported that Banp binds to the nucleotide sequence TCTCGCGAGA, called “Banp motif” and regulates transcription of essential metabolic genes in human and mouse (Grand et al., 2021). To identify direct targets of Banp in zebrafish, we examined whether genes whose mRNA level or chromatin accessibility is decreased in banpmutants carry Banp motif in their promoters. We first searched for enriched motifs in these genes using motif discovery software called HOMER with ATAC-seq data. Consistent with previous studies on human and mouse Banp (Grand et al., 2021), we found that TCTCGCGAGA is the most enriched motif in the closed chromatin region of banp mutants. Indeed, RNA-seq yielded 31 candidates of direct target genes from 81 downregulated genes, whereas ATAC-seq yielded 72 candidates, with 26 target genes consistent with both. So, these 26 candidates are the most likely to be target genes of Banp (Fig. 7A). Among them, we identified 8 cell-cycle regulators including cenpt, ncapg, and wrnip1 (Fig. 7B). We confirmed that wrnip1 mRNA is expressed in retinal progenitor cells in zebrafish and its mRNA expression is significantly reduced in banp mutants (Fig. 7CD). Wrnip1 promotes fork restart after replication stress, so these data can explain why cells with DNA replication stalling is increased in banp mutants.
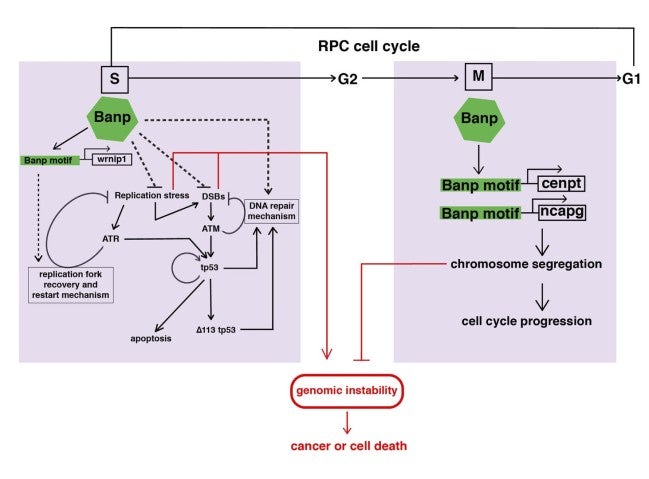
In summary, Banp serves essential functions in cell-cycle regulation in zebrafish retina (Fig. 8). During S phase, Banp suppresses DNA replication stress by activating transcription of a DNA replication stress regulator, wrnip1, and promotes DNA damage repair in concert with p53-dependent DNA damage response. In M phase, Banp promotes mitosis by activating transcription of two chromosomal regulators, cenpt and ncapg, most likely via their Banp motifs. Thus, Banp regulates progression in S phase and M phase through transcription control of distinct sets of cell-cycle regulators.
3.2 Machanism of photoreceoptor degeneration
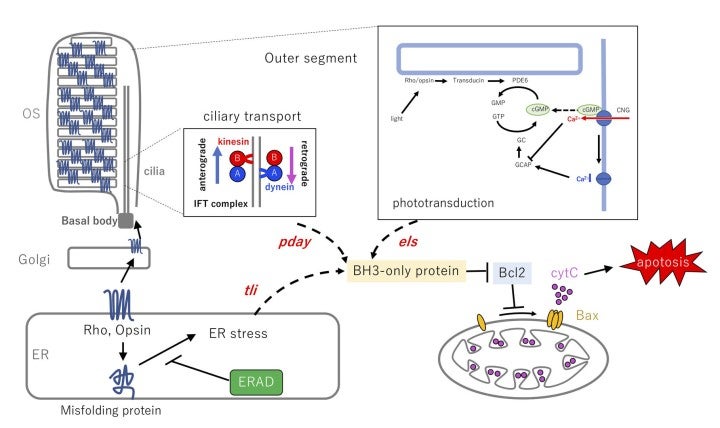
Apoptosis is observed in developing tissues and is believed to remove abnormal cells. Although apoptosis is important for establishment of proper neural circuits by eliminating abnormally differentiated neurons, it is unclear how differentiating cells monitor their own abnormality, and how the threshold at which apoptosis is induced is determined. Photoreceptors provide a useful model for studying such a surveillance mechanism of neuronal development and homeostasis, because there are many hereditary retinal diseases in humans associated with photoreceptor degeneration. Although 281 genes linked to hereditary retinal diseases have already been identified, these genes encode diverse functions, including phototransduction, retinol metabolism, and intracellular protein transport (see the homepage of the Retinal Information Network at http://www.sph.uth.tmc.edu/Retnet/). To answer how photoreceptors monitor their differentiation status and homeostasis, and what kinds of molecular network determine the choice between cell survival and cell death in photoreceptors, we have investigated zebrafish photoreceptor degeneration mutants. Previous zebrafish mutagenesis identified three zebrafish mutants namely els, tli and pday, in which photoreceptor degeneration is triggered by defects in phototransduction, ER quality control, and intraflagellar transport via the primary cilium (Nishiwaki et al., 2008) (Fig. 9). In FY2022, we focused on these photoreceptor degeneration mutants and investigated how defects in phototransduction, ER quality control, primary cilium-mediated protein transport cause photoreceptor degeneration.
3.3 The role of microglia in retinal development and degeneration
Microglia are brain-resident immune cells, originally derived from mesoderm-derivative tissue or the hematopoietic stem cell-lineage, that migrate into brain, and patrol within the brain throughout life. Microglia are thought to eliminate dead or dying neurons to prevent inflammation. Interestingly, microglia-mediated inflammation is required for neuronal regeneration in response to traumatic brain injury in zebrafish (Kyritsis et al., 2012), suggesting that microglia interact with neural stem cells and promote neural stem cells to initiate regeneration program in response to brain damage. Thus, it is important to understand the role of microglia in neuronal degeneration and regeneration.
3.3.1 Mechanism underlying microglia colonization into the developing retina
In FY2021, we examined normal developmental profiles of microglial precursors and their colonization mechanism to the zebrafish retina. It was reported that microglial precursors are initially generated in lateral plate mesoderm, but that they move to yolk, and enter the brain, including the eyes during zebrafish development (Herbomel et al., 2001). After microglial precursors enter the brain, they seem to become mature and start to express markers of functional microglia. However, it is unknown what kinds of guidance cues enable microglial precursors to move from yolk into developing retina, and how microglial precursors become functionally mature microglia after the enrty of brain.
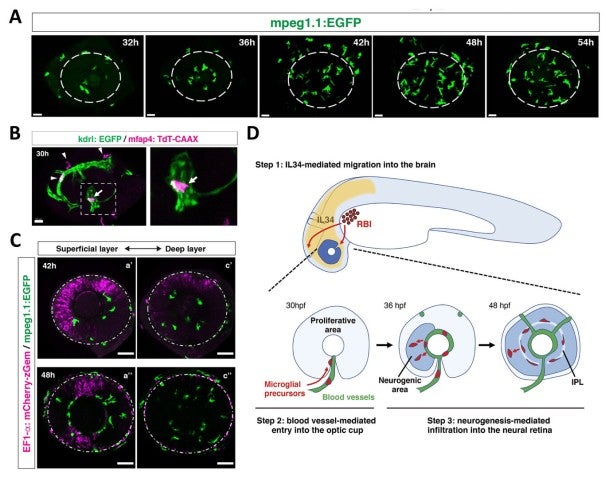
First, to elucidate guidance mechanism that promotes microglial precursors’ colonization into the brain, we conducted time-lapse observation using zebrafish mpeg1:GFP or mfap4:tdTomato transgenic lines, which specifically visualizes microglial precursors in zebrafish (Ellett et al., 2011; Walton et al., 2015). We found that microglial precursors progressively enter the optic cup from 24 to 54 hpf through the ventral optic fissure (Fig. 10A). Intraocular blood vessels initially form from the ventral fissure and then expand near the lens, suggesting an overlap with the microglial precursors’ colonization route. Indeed, microglial precursors are associated with surfaces of intraocular blood vessels. Furthermore, blockade of blood vessel formation reduced the number of microglial precursors in the retina (Fig. 10B), suggesting that blood vessel plays a role in guidance scaffold. We also found that microglial precursors enter the neural retina mainly through the retinal neurogenic area (Fig. 10C). These data suggest that neurogenesis functions as a gate that allows microglial precursors to move into the neural retina. It was reported that brain-derived interleukin 34 (IL34) and its receptor, colony-stimulating factor 1 receptor a (csf1ra) regulate colonization of microglial precursors in zebrafish brain and eye (Wu et al., 2018). We confirmed that IL34 is required for colonization of microglial precursors into the retina. Thus, our findings suggest a step-by-step guidance mechanism by which microglial precursors move from yolk into the optic cup and colonize the neural retina (Fig. 10D).
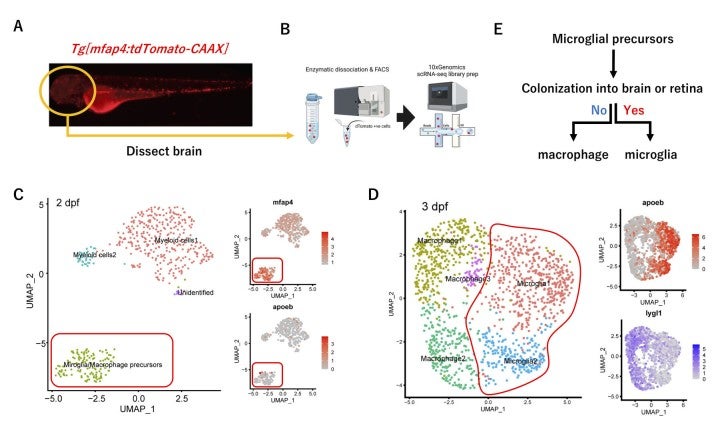
In FY2022, we investigated the second question on maturation mechanism of microglia. To understand maturation process of microglial precursors into functional microglia, we used zebrafish transgenic lineTg[mfap4:tdTomato-CAAX], which visualizes microglia/macrophage precursors, microglia and macrophage (Fig. 11A). We collected mfap4:tdTomato-CAAX-positive cells from zebrafish embryonic heads at 2, 3 and 5 dpf using FACS, applied single cell RNA-seq analysis (Fig. 11B), and conducted clustering of their dataset separately, using the Seurat package. We found that 2 dpf, mfap4-expressing cells form only one cluster (Fig. 11C). This single cluster expresses both microglial and macrophage markers ccl34b.1 and lygl1, but importantly does not express a mature microglial marker apoeb. Thus, it is very likely that 2 dpf mfap4-expressing cells are macrophage/microglial precursors. At 3 dpf, mfap4-expressing cells are separated into 5 clusters (Fig. 11D). Among them, only two clusters express apoeb, suggesting that mature microglia appear by 3 dpf. Importantly, in these apoeb-positive clusters, the macrophage marker lygl1 expression is downregulated and another microglial marker ccl34b.1 expression is upregulated. On the other hand, lygl1 expression is maintained and ccl34b.1 expression is downregulated in other three apoeb-negative clusters. Thus, fate determination of microglia and macrophage proceeds at 3 dpf. At 5 dpf, cell fate for mature microglia and macrophages is more clearly segregated in mfap4-expressing cells. Thus, these data revealed that mfap4-expressing cells at 2 dpf are reasonably classified as macrophage/microglial precursors, and a fraction of them differentiate into mature microglia after 3 dpf.
Next, we generated double transgenic line Tg[mfap4:tdTomato-CAAX; ccl34b.1:EGFP], by which we can distinct microglia and macrophage as mfap4+; ccl34b+ cells and mfap4+; ccl34b– cells, respectively at 5 dpf. We confirm that almost all of mfap4+; ccl34b+ cells are located inside the brain; however, a majority of mfap4+; ccl34b–cells are located outside the brain. These data suggest that only microglial/macrophage precursors that enter the brain can differentiate into mature microglia (Fig. 11E). This is consistent with a classic and pioneer work indicating that early macrophages outside the brain never express Apoe (Herbomel et al., 2001). Our findings provide in vivo evidence on how microglial precursors differentiate into mature microglia.
3.3.2 The role of microglia in neural stem cell activation in response to trauma brain injury
In mammals, brain damage induces acute inflammation, microglial activation, and reactive astrogliosis, leading to scar formation, which limits spread of damage into unaffected CNS area. However, significant regeneration does not occur. In contrast, stab injuries to the zebrafish telencephalon cause an acute inflammatory response and reactive gliosis; however, neither persistent gliosis, fibrotic scar formation nor chronic inflammation occurs. Rather, radial glial cells (RGs) upregulate cell proliferation at 2-3 days post-lesion (dpl), and subsequently promote neurogenesis, which leads to rebuild neural circuits of damaged tissue. So, RGs show a remarkable ability to regenerate damaged neural tissue by re-initiating cell proliferation to produce neural precursors to rebuild the lost neural circuit. However, it is not fully understood how RGs respond to brain damage to initiate this regenerative response.
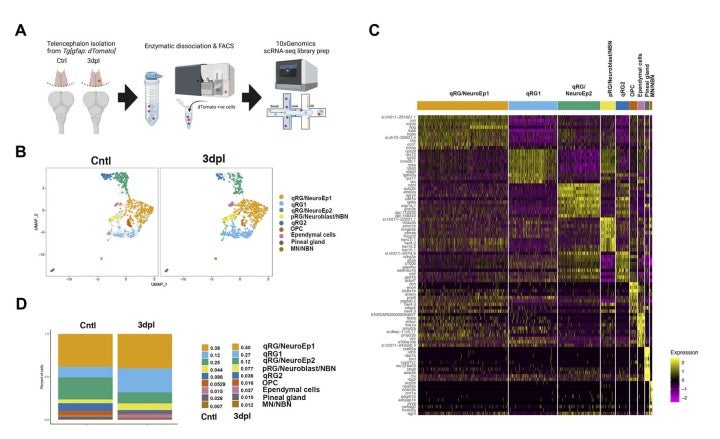
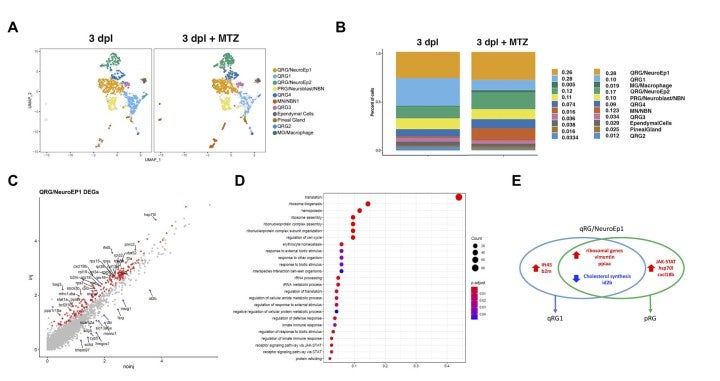
To understand how RGs respond to brain damage to initiate a regenerative response, we applied single-cell transcriptomics to RGs in zebrafish adult telencephalon using the zebrafish transgenic line Tg[GFAP:dTomato] (Fig. 12A). We identified five RG subtypes, which are classified into four quiescent RGs (qRG) and one proliferating RG (pRG) (Fig. 12B). The four qRGs express distinct subsets of qRG markers, suggesting heterogeneity of qRGs in adult zebrafish brain (Fig. 12C). Interestingly, one qRG subtype shows high expression of ribosomal proteins, and its fraction increases in response to brain damage (Fig. 12D). Consistently, the mTOR pathway is activated in RGs near the injury site.
Next, to investigate the role of microglia in regenerative responses of RGs, we eliminated microglia. We confirmed that microglial elimination suppresses damage-induced regenerative responses of RGs. Interestingly, our scRNA-seq analysis revealed that the fraction of ribosomal expression-enriched qRGs is markedly decreased in microglia-depleted injury brain, indicating that the transition of ribosomal expression-enriched qRGs depends on microglia function (Fig. 13AB). Our pseudo-time analysis suggests that putative dormant RGs produce the ribosomal expression-enriched qRGs through activation of ribosomal genesis as well as suppression of cholesterol biogenesis, and pRGs through activation of the JAK/STAT pathway (Fig. 13CDE). Our findings reveal heterogeneity of qRGs in adult zebrafish brain and their dynamic regenerative responses to brain damage.
3.4 Lens fiber differentiation
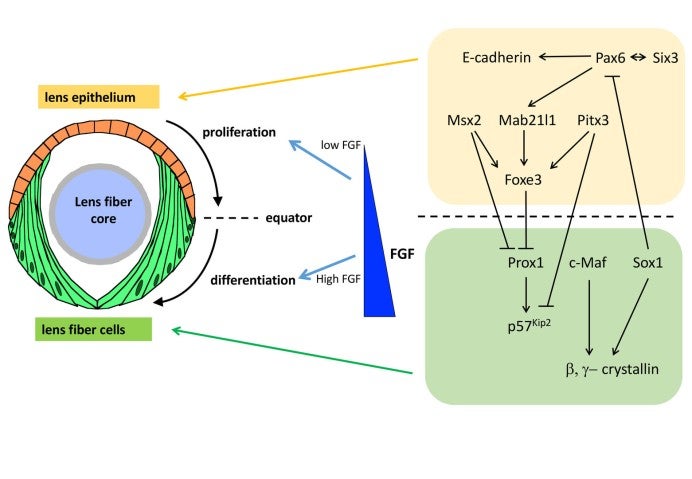
The lens is an intraocular organ that focuses visual images on retinal photoreceptors. The lens consists of two cell types: lens epithelial cells and lens fiber cells. Lens epithelium convers the anterior half of the lens fiber core (Lovicu and McAvoy, 2005; Mochizuki and Masai, 2014). At the lens equator, epithelial cells start to differentiate into lens fiber cells, which elongate and pile up to cover the old lens fiber core. Thus, the lens provides a good model for studying spatiotemporal coordination of cell differentiation and morphogenesis (Fig. 14). In vertebrate lens, FGF is secreted from the retina and is believed to form a low-high gradient along the anterior-posterior axis of the lens. It was reported that FGF promotes cell proliferation of lens epithelial cells at low doses and lens fiber differentiation at high doses (McAvoy et al., 2917) (Fig. 14). These observations raised the “FGF gradient hypothesis”, in which FGF regulates multiple steps of lens fiber differentiation: low doses of FGF promote lens epithelial cell proliferation, whereas high doses of FGF induce lens fiber cell differentiation. Furthermore, previous studies revealed a network of transcription factors that coordinate lens epithelial cell proliferation in the anterior lens and lens fiber differentiation in the posterior lens (Fig. 14). These transcription factors are classified into two groups. In the anterior lens epithelium, Pax6 activates Mab21l1 and Foxe3 in concert with Pitx3 and Msx2, resulting in maintenance of E-cadherin expression and a proliferative state of lens epithelial cells. In the posterior lens fiber region, Prox1 activates cell-cycle inhibitor, p57kip2, to promote cell-cycle exit of lens epithelial cells at the equator. In addition, c-Maf and Sox1 activate expression of b- and g-cyrstallin to promote lens fiber maturation. These two groups of transcription factors mutually repress each other to maintain the boundary between the lens epithelium and lens fiber areas (Fig. 14). However, the mechanism that spatially regulates lens epithelial proliferation and lens fiber differentiation is still not fully understood.
3.4.1 Identification of FGF ligands that regulate lens fiber differentiation in zebrafish
Unlike other signaling pathways, FGF signaling is unique, because more than 20 types of FGF ligands are identified in vertebrates (Ornitz and Itoh, 2015), which contribute to their functional redundancy and robust developmental and morphogenetic processes (Leerberg et al., 2019). Although FGF is essential for lens development in zebrafish, it is still unknown what types of FGF ligands are involved in lens epithelial proliferation and lens fiber differentiation. To understand molecular mechanisms of lens fiber differentiation, it is necessary to identify FGFs that regulate lens development in zebrafish. I examined all the FGF genes expressed in zebrafish retina using RT-PCR and in situhybridization. Among the total 27 FGF ligands, Fgf3 and Fgf8a are predominantly expressed around the developing lens in zebrafish. Fgf3 is expressed in the ventral region of the retina, which is just beneath lens placode, from 15 somite stage to18 hpf. On the other hand, Fgf8a is expressed in newly differentiating retinal ganglion cells (RGCs), whose position is very close to the lens equator, after 36 hpf. To determine the roles of Fgf3 and Fgf8a, I applied their morpholino antisense oligos. In fgf3 morphants, expression of an FGF target gene, pea3, is suppressed in the lens at 18-24 hpf, resulting in small lenses at 34 hpf. However, pea3 was normally expressed at 18 -24 hpf in fgf8a morphant lens. So, Fgf3 is the major FGF ligand that regulates lens development, especially lens vesicle size by promoting lens placodal cell proliferation. On the other hand, at 38 hpf, pea3 was normally expressed in lens equatorial region in fgf3 morphants; however, pea3expression was suppressed in lens equatorial region of fgf8a morphants. Indeed, in fgf8a morphants, the entry point of lens fiber differentiation was more shifted posteriorly at 38 hpf and the number of newly differentiating lens fiber cells was significantly reduced at 48 hpf, resulting in defects in spherical lens shape formation. So, Fgf8a mainly regulates lens fiber differentiation. These observations imply that FGF3 and FGF8a differentially regulate lens development: FGF3 secreted from the ventral retina regulates lens epithelial cell proliferation in the early stage, whereas Fgf8a secreted from differentiating RGCs regulates lens fiber cell differentiation in the later stage.
3.4.2 The role of FGF receptor like 1 (FGFRL1) in lens fiber differentiation
To elucidate the mechanism underlying equator-specific onset of lens fiber differentiation, we focus on one of Fgfrl1 genes, Fgfrl1b, which is specifically expressed in lens epithelium. FGF receptor-like 1 (Fgfrl1) has three extracellular immunoglobulin domains, which bind to FGF ligands, but lacks an intracellular kinase domain. In mammals, proteolytic cleavage of Fgfrl1 releases its ectodomain, which subsequently binds FGF ligands, suggesting that Fgfrl1 functions as a decoy receptor. To inhibit the function of Fgfrl1b, we injected its morpholino antisense into wild-type embryos and examined lens fiber cell differentiation. In Fgfrl1b morphant, expression of an FGF target was elevated, suggesting that Fgfrl1b suppresses FGF signaling. Consistently, the expression of a lens fiber differentiation marker was enhanced. Ectopic differentiation also occurred in lens epithelium of Fgfrl1b morphant. Thus, Fgfrl1b suppresses FGF signaling to ensure equator-specific onset of lens fiber differentiation.
3.5 Zebrafish behavioral mechanism: The role of Mecp2 in fear/anxiety regulation in zebrafish brain
Rett syndrome is an autism spectrum disorder and one of the most common causes of mental retardation in women. Sporadic mutations on the transcription factor MeCP2 has been found as one of the most frequent causes for Rett syndrome in humans but how MeCP2 mutations cause of Rett syndrome is still largely unknown (Kyle et al. 2018). In recent years multiples models of Rett syndrome had been developed in mice and zebrafish (Pietri et al. 2013). Mice models show affected social behavior, but so far there are no good descriptions of how MeCP2 mutations affect the social behavior adult zebrafish. To elucidate the role of MeCP2 in behaviors, I examined behavioral phenotypes in zebrafish MeCP2 mutants. Interestingly, MeCP2 mutant adult zebrafish showed increased anxiety and fear response, as well as reduced thigmotaxis, which are reminiscent of the Rett syndrome phenotype, specifically high anxiety. On the other hand, they showed normal social preference. Next, we examined whether zebrafish MeCP2 mutants harbor defects in brain development. Our imaging analysis revealed that zebrafish MeCP2 mutants showed increased size in the DM region of the telencephalon, a region homologous to the amygdala in mammals and responsible for fear behaviors, compared with wild-type siblings. These data suggest that MeCP2 plays a crucial role in neural circuit formation responsible for fear response in telencephalon.
4. Publications
4.1 Journals (* corresponding authors)
- Babu, S., Takeuchi, Y. and Masai, I.* (2022) Banp regulates DNA damage response and chromosome segregation during the cell cycle in zebrafish retina. eLife 11:e74611. doi: 10.7554/eLife.74611.
- Kutsia, M., Takeuchi, Y., and Masai, I.* (2022) Single -cell transcriptome analysis reveals heterogeneity and a dynamic regenerative response of quiescent radial glia in adult zebrafish brain. bioRxiv 2022.07.27.501663. doi; https://dpi.org/10.1101/2022.07.27.501663.
4.2 Books and other one-time publications
Nothing to report
4.3 Oral and Poster Presentations
(Oral, International conference)
- Kutsia, M., Takeuchi, Y., and Masai, I. Single cell transcriptome analysis reveals heterogeneity and a dynamic regenerative response of quiescent radial glia in zebrafish adult brain. International Zebrafish Conference (IZFC2022), 22-26 June 2022. Montreal Canada (online).
- Takeuchi, Y., and Masai, I. Fgf3 and Fgf8a differentially regulate lens vesicle formation and lens fiber differentiation in zebrafish. ISER2023 (XXV Biennial Meeting of the International Society for Eye Research). Gold Coast, Australia, 19-23 February 2023.
- Sugiyama, Y. Lovicu, F., Robinson, M., and Masai, I. Fibroblast growth factor differentially regulates Rho/Rac activity to drive fiber cell elongation in the mammalian lens. ISER2023 (XXV Biennial Meeting of the International Society for Eye Research). Gold Coast, Australia, 19-23 February 2023.
(Poster, international conferences)
- Rodriguez, L. C., Izawa, T., and Masai, I. Zebrafish MeCP2 mutation produce behavioral alterations reminiscent of Rett Syndrome. International Zebrafish Conference (IZFC2022), 22-26 June 2022. Montreal Canada (online).
- Ahmed, M., Kojima, Y., and Masai, I. Strip1 coordinates retinal neural circuit formation by promoting retinal ganglion cell survival during development. International Zebrafish Conference (IZFC2022), 22-26 June 2022. Montreal Canada (online).
- Babu, S., Takeuchi, Y., and Masai, I. Banp is an essential transcription factor that regulates DNA damage response and chromosome segregation during the cell cycle. International Zebrafish Conference (IZFC2022), 22-26 June 2022. Montreal Canada (online).
- Chiang, H., Nishiwaki, Y., and Masai, I. Male germ cell-associated kinase is essential for zebrafish photoreceptor ciliogenesis and cell survival. EMBO Workshop “Cilia 2022”, 4-7 October 2022, Cologne, Germany (online).
(Invited talks)
- Ichiro Masai, Genetic dissection of retinal neural circuit formation in zebrafish. A Recipe for Scientific Synergy -Series 2- OIST-Osaka University Joint Workshop (online), 28 April 2022.
- Ichiro Masai, Mechanism of microglia colonization of the retina and its role in neuronal degeneration. Retinal Research Meeting (RRM), Tokyo, 26 November 2022.
- Ichiro Masai, Genetic dissection of neural circuit formation in zebrafish retina. OIST workshop “Nervous system assembly”, 6-9 March 2023.
(Oral, domestic conferences)
- Ahmed, M., Kojima, Y., and Masai, I. Strip1 coordinates retinal neural circuit formation by promoting retinal ganglion cell survival during development. Annual Meeting of Japanese society of Developmental Biologists. 31 May-3 June 2022, Kanazawa, Japan.
- Ravishankar, D. Takeuchi, Y. and Masai, M. Roles for microglia in chronic photoreceptor degeneration and associated regeneration caused by pde6c dysfunction in zebrafish. Annual Meeting of Japanese society of Developmental Biologists. 31 May-3 June 2022, Kanazawa, Japan.
- Rodriguez, L. C., Izawa, T., and Masai, I. Zebrafish MeCP2 mutation produce behavioral alterations reminiscent of Rett Syndrome. Annual Meeting of Japanese society of Developmental Biologists. 31 May-3 June 2022, Kanazawa, Japan.
- Takeuchi, Y., Kojima,Y. and Masai, I. Fgf8a secreted from differentiating retinal ganglion cells regulates lens fiber differentiation in zebrafish. Annual Meeting of Japanese society of Developmental Biologists. 31 May-3 June 2022, Kanazawa, Japan.
- Rodriguez, L. C., Izawa, T. and Masai, I. Zebrafish MeCP2 mutation produce behavioral alterations reminiscent of Rett Syndrome. The 28th Japanese Zebrafish and Medaka Meeting, 1-2 September 2022, Osaka, Japan.
(Poster, domestic conferences)
- Masaki, T., Takeuchi, Y. and Masai, I. Investigating the Anti-apoptotic Mechanisms in the Retinal Ciliary Marginal Zone. Annual Meeting of Japanese society of Developmental Biologists. 31 May-3 June 2022, Kanazawa, Japan.
- Nishiwaki, Y., Fujiwara, M., and Masai, I. Drp1 is required for photoreceptor apoptosis in response to vesicular fusion defects. Annual Meeting of Japanese society of Developmental Biologists. 31 May-3 June 2022, Kanazawa, Japan.
- Nishiwaki, Y., Fujiwara, M., and Masai, I. Drp1 is required for photoreceptor apoptosis in response to vesicular fusion defects. The 28th Japanese Zebrafish and Medaka Meeting, 1-2 September 2022, Osaka, Japan.
- Nishiwaki, Y., and Masai, I. Photoreceptor degeneration in zebrafish pde6c mutants depends on elevation of intracellular Ca2+ through CNG3. The 45th Annual Meeting of Japanese Society of Molecular Biology, Makuhari, Ciba, Japan. 30thNovember – 2nd December 2022.
5. Intellectual Property Rights and Other Specific Achievements
5.1 The Category or Type of Funding, like External Funding, Awards, etc.
5.1.1 Funding
KAKENHI (grants from the Ministry of Education, Science and Sport/JSPS)
- PI name: Yuko Nishiwaki
- Title: Mechanism of Drp1-mediated formation of endoplasmic reticulum-mitochondria contact site and its role in apoptosis induction
- Category: Grant-in-Aid for Scientific Research (C)
- Period: FY2021-FY2023
- PI name: Yuki Siguyama
- Title: Analysis of mechanical interaction between basal membrane and lens fiber cells during lens morphogenesis
- Category: Grant-in-Aid for Scientific Research (C)
- Period: FY2021-FY2023 (Completed by FY2021)
- PI name: Wei-Chieh Chiang
- Title: The mechanism of endoplasmic reticulum proteostasis and proteotoxicity in retinal degeneration.
- Category: Grant-in-Aid for Scientific Research (C)
- Period: FY2020-FY2023
- PI name: Yuki Takeuchi
- Title: Identification of FGF ligands involved in the regulation of lens fiber differentiation using zebrafish model.
- Category: Grant-in-Aid for Scientific Research (C)
- Period: FY2022-FY2025
- PI name: Darshini Ravishankar
- Title: Temporal profiling of photoreceptor degeneration in zebrafish PDE6c mutants and a potential role of microglia
- Category: JSPS DC2
- Period: FY2022-FY2023
6. Meetings and Events
6.1 Seminar Title in Full
- Lecturer: Dr. Yohei Ogawa (Department of Pathology and Immunology, Washington University School of Medicine, US)
- Title: Conservation and divergence of retinal cell types during vertebrate evolution
- OIST campus (online)
- Date: 29 June 2022
6.2 OIST workshop
None
7. Other
Nothing to report.