




Developmental Neurobiology Unit
Associate Professor Ichiro Masai
Research Theme: Mechanisms underlying retinal and lens development

Abstract
The vertebrate neural retina is derived from the ventral region of the forebrain. In this region, six classes of neurons differentiate and form the neural circuit underlying visual transduction. Thus, the retina provides an excellent model for studying cell differentiation and neural circuit formation in the vertebrate brain. Furthermore, more than one hundred hereditary retinal diseases causing photoreceptor degeneration have been identified in humans. Understanding the pathological processes of photoreceptor degeneration is an important issue from a medical perspective. We are currently investigating the mechanisms underlying retinal cell differentiation and photoreceptor degeneration, using zebrafish as an animal model. The results have revealed that five signaling molecules, including Histone deacetylase 1 (Hdac1), regulate retinal neurogenesis in zebrafish. Regulatory factors that modulate the Hdac1 activity during retinal neurogenesis have been examined. Furthermore, to examine cell-cycle regulation in retinal neurogenesis with high accuracy, we visualized cell-cycle phases differentially with fluorescent proteins using zebrafish fluorescent ubiquitination-based cell cycle indicator (Fucci). We applied this Fucci technique to zebrafish lens epithelium and revealed a spatial regulation of cell proliferation and migration. In the project on photoreceptor degeneration, we have identified zebrafish mutants showing photoreceptor degeneration. In 2012, we revealed that a zebrafish b-SNAP mutation causes photoreceptor apoptosis, which depends on a BH3-only SNARE, BNip1. Because b-SNAP regulates the fusion of transport vesicles to intracellular membrane organelles, this finding suggests that BNip1 mediates photoreceptor apoptosis in response to vesicular fusion defects.
1. Staff
- Dr. Yuko Nishiwaki, Researcher
- Dr. Toshiaki Mochizuki, Researcher
- Dr. Junichi Kawada, Researcher
- Dr. Maria Iribarne, Researcher
- Ms. Asuka Yoshizawa, Technician
- Mr. Yutaka Kojima, Technician
- Mr. Shohei Nakamura, Technician
- Ms. Eri Oguri (temporal worker), Technician
- Ms. Yayoi Tomoyose, Research Assistant
- Ms. Ayako Nameki, Research Assistant
- Ms. Kazumi Toguchi, Research Assistant
- Ms. Yuko Yamazato, Research Assitant
- Dr. Mariko Kinoshita-Kawada, Research Assistant
- Ms Ayako Gima, Research Administrator/Secretary
2. Collaborations
- Theme: In vivo functional analysis of hypoxia response genes using the zebrafish retina
- Type of collaboration: Joint research
- Researchers:
- Professor, Masayuki Matsushita, Ryukyu University
3. Activities and Findings
3.1 Mechanism regulating retinal neurogenesis in zebrafish
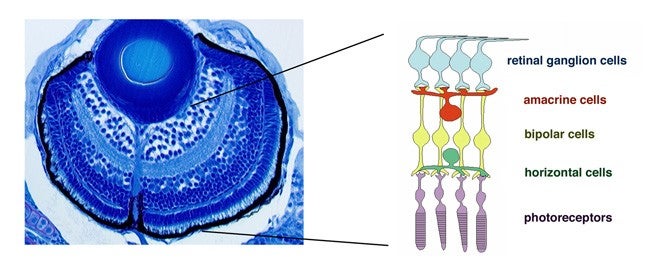
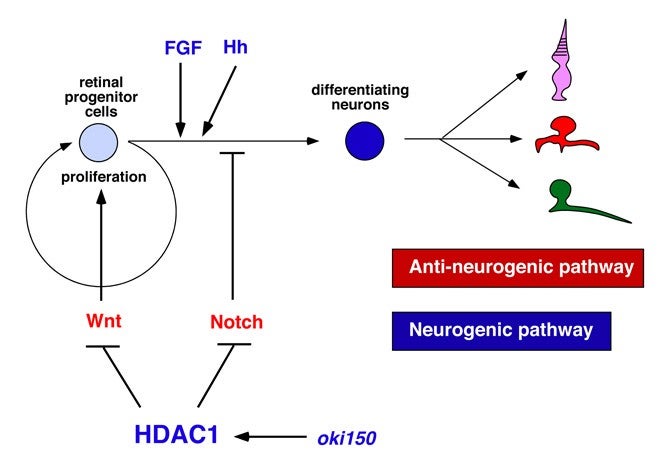
The vertebrate retina is initially specified in the anterior neural plate and evaginates from the ventral region of the forebrain as the optic cup. In this region, six major classes of retinal neurons and one class of glial cells differentiate to form the neural circuit that mediates phototransduction and visual processing (Fig. 1). Thus, the retina provides an excellent model for studying the mechanisms underlying cell differentiation and neural circuit formation in the developing brain. In the developing zebrafish retina, neurogenesis is initiated at a few cells adjacent to the optic stalk and progresses to the entire neural retina. This pattern of retinal neurogenesis provides a good model for studying the mechanisms regulating the spatial and the temporal pattern of neurogenesis in the nervous system. Previous studies, including research in our lab, suggest that at least five families of signaling molecules regulate the pattern of retinal neurogenesis in zebrafish (Fig. 2). Fibroblast growth factors (Fgfs) are expressed in the optic stalk and are required for the initial induction of retinal neurogenesis in zebrafish. The Hedgehog (Hh) signaling pathway is important for the progression of retinal neurogenesis in zebrafish. Activation of the Wnt signaling pathway promotes cell proliferation in the zebrafish retina, and activation of the Notch pathway inhibits retinal neurogenesis. Previously, we identified a zebrafish histone deacetylase 1 (hadc1) mutant, in which retinal progenitor cells fail to exit from the cell cycle and continue to proliferate. HDAC1 promotes retinal neurogenesis by suppressing both the Wnt and Notch signaling pathways. HDAC1 is recruited to several transcription repressor and corepressor complexes as well as those in the Wnt and Notch pathways. To elucidate the HDAC1-dependent regulation of retinal neurogenesis, we have identified a new mutation, oki150, that modifies the neurogenic defects in heterozygous hdac1 mutants. In 2011, using polymorphic markers, we mapped the oki150 mutational locus on zebrafish chromosome 20. We currently clone the oki150 mutation.
3.2 Visualization of cell-cycle phases with fluorescent proteins using Fucci
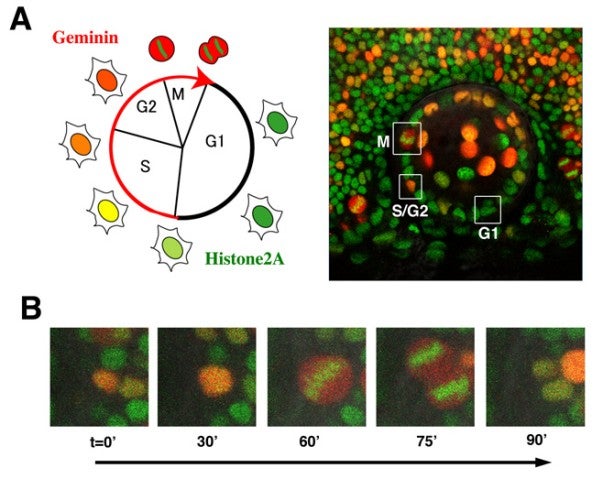
The first step of neurogenesis is the exit from the cell cycle. Previously we showed that the earliest neurogenic marker, ath5 (also designated as atoh7), is initiated in the G2 phase in retinal progenitor cells just prior to the final cell division, which generates two neuronal daughter cells. Thus, it is very likely that the commitment to the retinal neuron is associated with cell-cycle progression. To accurately understand the relationship between cell proliferation and cell differentiation, we visualized cell-cycle phases differentially with fluorescent proteins, using the zebrafish Fucci (fluorescent ubiquitination-based cell cycle indicator) (Sugiyama et al., 2009, Proc Natl Acad Sci USA 106, 20812–20817). In the Fucci system, the cell-cycle dependent expression of two cell-cycle regulators, Geminin and Cdt1, are visualized with fluorescent proteins fused to their regulatory peptide sequences. We generated the zebrafish transgenic line carrying genes encoding mCherry-tagged Geminin and green fluorescent protein (GFP)-tagged Histone 2A. In this transgenic fish, we were able to clearly distinguish three different phases in the developing retina as well as the lens; the G1, S-G2, and M phases (Fig. 3A). Furthermore, we were able to observe chromosome condensation in the M phase (Fig. 3B) and measure the orientation of cell division from in vivo time-lapse imaging data.
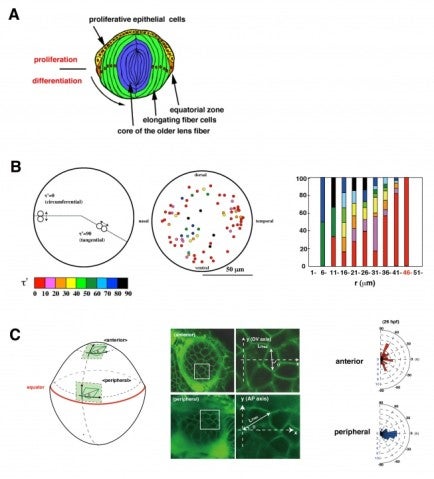
In 2012, using this transgenic fish, we observed the cell-cycle progression, cell division, and cell migration in the zebrafish lens epithelium using time-lapse techniques, because the lens epithelium is more easily accessed than the neural retina (Fig. 4A). We found that cell proliferation was highly active in a marginal zone just anterior to the equator in the lens epithelium. Furthermore, cell-division orientation was biased along the circumferential axis in this proliferating zone (Fig. 4B). These data suggest that cell proliferation and cell division are dynamically regulated in the zebrafish lens epithelium. In many developing tissues, it has been reported that cells tend to divide along their longest axis. This is referred to as “the Hertwig rule”. We found that the longest axis was also circumferentially biased in the proliferating zone in the zebrafish lens epithelium (Fig. 4C), suggesting that lens epithelial cells follow the Hertwig rule. These observations suggest that apical cell geometry correlates with cell division orientation in the proliferating zone of the lens epithelium. Since cell geometry depends on cell adhesion and a cell adhesion molecule, E-cadherin, is expressed in the lens epithelium, we are examining epithelial cell behaviors in zebrafish E-cadherin mutant.
3.3 Mechanism underlying axonal guidance crossing the midline
Visual information processed in the retina is transmitted from retinal ganglion cells to the second visual center, called the optic tectum, through retinal axons. In lower vertebrates including fish, all retinal axons project to the contra-lateral side of optic tectum and no ipsi-lateral projection occurs, resulting in the formation of the optic chiasm, where retinal axons from the left and right eyes cross without combining. Our previous study showed that retinal axons are misrouted to the ipsi-lateral side of the optic in zebrafish N-cadherin mutants, suggesting a role of cell adhesion in retinal axon guidance at the midline. However, the mechanism underlying retinal axon guidance in crossing on the midline remains to be elucidated.
The molecular mechanism underlying axon guidance at the midline has been studied using commissural neurons in the vertebrate hindbrain and spinal cord as well as in the Drosophila embryonic central nervous system (CNS). It is generally accepted that pre-crossing axons are attracted by a guidance cue emanating from the midline tissue, Netrin, and that the midline tissue expresses not only Netrin but also a repellent cue, Slit. A recent model proposes that pre-crossing axons are insensitive to Slit and only respond to Netrin, resulting in the attraction of their growth cone to the midline. When their growth cone passes over the midline, the sensitivity to Slit is increased, which enables post-crossing axons to leave from the midline. It has been reported that the expression level of a receptor of Slit, namely Roundabout (Robo), is very low in pre-crossing axons but increased in post-crossing axons in Drosophila CNS. In addition, it was reported that one of the Robo family proteins, Robo3, is exclusively expressed in the pre-crossing axons and functions as dominant-negative repressor of Robo1, which is thought to mediate repulsive response instructed by Slit. However, the way in which the responsiveness of Slit is differentially regulated in pre- and post-crossing axons remains unclear.
We have established an in vitro culture assay system using commissural axons that reflects the acquisition of Slit responsiveness in post-crossing axons. Using this assay system, we showed that a Robo1-associating deubiquitinase, USP33, is required for the acquisition of Slit responsiveness and crossing of commissural axons over the midline. From 2011 to 2012, we revealed that the endocytic trafficking of Robo1 modulates its own expression level on axons as well as the sensitivity of axons to Slit. These findings suggest a new mechanism that modulates axonal responsiveness to guidance cues in crossing the midline of the CNS. We are confirming the role of the endocytic trafficking in axon crossing the midline in vivo using mouse and chick neural tube.
3.4 Mechanisms underlying photoreceptor degeneration
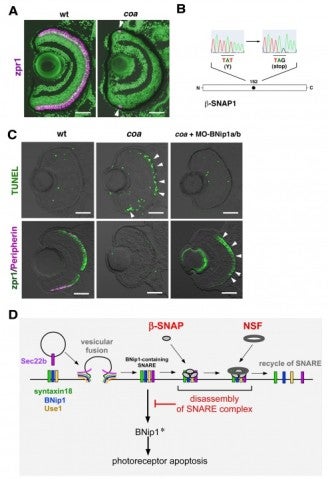
Photoreceptors are highly specialized neurons that efficiently detect light stimuli. To date, more than a hundred genes associated with inherited photoreceptor degeneration have been identified in humans (see the homepage of the Retinal Information Network at http://www.sph.uth.tmc.edu/Retnet/). However, the mechanisms underlying the pathological processes involved in photoreceptor degeneration also remain to be elucidated. To determine the mechanisms underlying photoreceptor degeneration, we have screened zebrafish mutants with dysfunctional visual behavior by testing the optokinetic response. We identified several zebrafish mutants, in which photoreceptors differentiate but later degenerate. In 2010, we reported that one of these mutants carried a genetic mutation in cGMP-phosphodiesterase 6c (PDE6c), a mediator molecule of cone-specific phototransduction, suggesting a link between achromatopsia and progressive cone dystrophy. In 2012, we examined another mutant, coa, whose photoreceptors rapidly undergo apoptosis before maturation (Fig. 5A). We found that the coa mutant gene encodes b-soluble N-ethylmaleimide-sensitive factor attachment protein (b-SNAP) (Fig. 5B). b-SNAP regulates the fusion of transport vesicles to intracellular membrane organelles. These observations suggest that vesicular transport defects cause photoreceptor apoptosis in zebrafish.
Mitochondria-dependent apoptosis is promoted by proapoptotic Bcl2 family proteins, Bax, and inhibited by anti-apoptotic Bcl2 family proteins, Bcl2. BH3-only proteins promote apoptosis by modulating the balance between pro-apoptotic and anti-apoptotic Bcl2 proteins. We searched which BH3-only proteins are involved in the coa-mediated photoreceptor apoptosis and found that a BH3-only SNARE, BNip1, mediates the coa-mediated photoreceptor apoptosis (Fig. 5C). BNip1 is a component of the syntaxin 18 SNARE complex, and the syntaxin 18 SNARE complex regulates retrograde transport from the Golgi apparatus to the ER. b-SNAP promotes the recycle of vesicular fusion machinery, SNARE, by disassembling the cis-SNARE complex generated by vesicular fusion. Thus, it is likely that the syntaxin 18 cis-SNARE complex is accumulated in the coa mutant. We found that failed disassembly of the syntaxin 18 cis-SNARE complex activates the BNip1-dependent photoreceptor apoptosis in the coa mutant. These data suggest that the sytaxin 18 cis-SNARE complex monitors vesicular fusion competence and that BNip1 transforms vesicular fusion defects into apoptosis (Fig. 5D). Our findings indicate a novel role for BNip1in linking vesicular fusion competence and photoreceptor apoptosis.
4. Publications
4.1 Journals
-
Nishiwaki, Y., Yoshizawa, A., Kojima, Y., Oguri, E., Nakamura, S., Suzuki, S., Yuasa-Kawada, J., Kinoshita-Kawada, M., Mochizuki, T. and Masai, I. (2013). The BH3-only SNARE BNip1 mediates photoreceptor apoptosis in response to vesicular fusion defects. Dev. Cell 25, 374 – 387.
4.2 Books and other one-time publications
Nothing to report
4.3 Oral and Poster Presentations
Oral, International conference
- Nishiwaki, Y., Yoshizawa, A., Kojima, Y., Oguri, E., Nakamura, S., Suzuki, S., Yuasa-Kawada, J., Kinoshita-Kawada, M., Mochizuki, T. and Masai, I. Genetic mutation of beta-SNAP causes photoreceptor degeneration through the activation of the BH3-only SNARE, BNip1. 10th International Conference Zebrafish Development and Genetics, Madison, USA, 20–24 June 2012.
- Mochizuki, T., Suzuki, S., Sakaue-Sawano, A., Miyawaki, A. and Masai, I. E-cadherin controls apical cell geometry and orientation of cell division in the zebrafish lens epithelium. 10th International Conference Zebrafish Development and Genetics, Madison, USA, 20–24 June 2012.
Poster, international conferences
- Nishiwaki, Y., Yoshizawa, A., Kojima, Y., Oguri, E., Nakamura, S., Suzuki, S., Yuasa-Kawada, J., Kinoshita-Kawada, M., Mochizuki, T. and Masai, I. Genetic mutation of beta-SNAP causes photoreceptor degeneration through the activation of the BH3-only SNARE, BNip1. APDBC Taiwan 2012 Asia-Pacific Developmental Biology Conference 2012, Taipei Innovation City Convention Center, Taipei, Taiwan, 5–8 October 2012.
- Mochizuki, T., Suzuki, S., Sakaue-Sawano, A., Miyawaki, A. and Masai, I. E-cadherin controls apical cell geometry and orientation of cell division in the zebrafish lens epithelium. APDBC Taiwan 2012 Asia-Pacific Developmental Biology Conference 2012, Taipei Innovation City Convention Center, Taipei, Taiwan, 5–8 October 2012.
- Nishiwaki, Y., Yoshizawa, A., Kojima, Y., Oguri, E., Nakamura, S., Suzuki, S., Yuasa-Kawada, J., Kinoshita-Kawada, M., Mochizuki, T. and Masai, I. The BH3-only SNARE of syntaxin18, BNip1, mediates photoreceptor apoptosis in response to vesicular fusion defects. Gordon research conference – visual system development. Colby-Sawyer College, New London, NH. 19–24 August 2012
Invited talks
- Mochizuki, T. and Masai, I. Spatial regulation of cell proliferation and differentiation in the zebrafish lens. The 46th Annual Meeting of the Japanese Society of Developmental Biologists Matsue, Shimane, Japan. 28–31 May 2013.
- Masai, I. Mechanism underlying lens differentiation through the ubiquitin-proteasome system. The joint meeting of the 27th annual meeting of the Japanese society of cataract and refractive surgery and the 51st annual meeting of the Japanese society of cataract research. Tokyo, Japan. 15–17 June 2012.
- Masai, I. Mechanism underlying lens differentiation using zebrafish mutants. Wakayama Medical University, 15 June 2012.
Oral, domestic conferences
- Nishiwaki, Y., Yoshizawa, A., Kojima, Y., Oguri, E., Nakamura, S., Suzuki, S., Yuasa-Kawada, J., Kinoshita-Kawada, M., Mochizuki, T. and Masai, I. The BH3-only SNARE of syntaxin18, BNip1, is a mediator of photoreceptor apoptosis in response to vesicular fusion defects. The 35th annual meeting of the Molecular Biology Society of Japan. Fukuoka, Japan. 11–14 December 2012.
- Nishiwaki, Y., Yoshizawa, A., Kojima, Y., Oguri, E., Nakamura, S., Suzuki, S., Yuasa-Kawada, J., Kinoshita-Kawada, M., Mochizuki, T. and Masai, I. Genetic mutation of beta-SNAP causes photoreceptor degeneration through the activation of the BH3-only SNARE, BNip1.The 18th Japanese Medaka and zebrafish meeting. Kyoto, Japan, 22–23 September 2012.
- Nishiwaki, Y., Yoshizawa, A., Kojima, Y., Oguri, E., Nakamura, S., Suzuki, S., Yuasa-Kawada, J., Kinoshita-Kawada, M., Mochizuki, T. and Masai, I. The BH3-only SNARE of syntaxin18, BNip1, is a mediator of photoreceptor apoptosis in response to vesicular fusion defects. Neuroscience 2012. The 35th annual meeting of the Japan Neuroscience Society. Nagoya, Japan. 18–21 September 2012.
- Nishiwaki, Y., Yoshizawa, A., Kojima, Y., Oguri, E., Nakamura, S., Suzuki, S., Yuasa-Kawada, J., Kinoshita-Kawada, M., Mochizuki, T. and Masai, I. Genetic mutation of beta-SNAP causes photoreceptor degeneration through the activation of the BH3-only SNARE, BNip1. Joint meeting of the 45th annual meeting of the Japanese society of Developmental Biologists and the 64th annual meeting of the Japanese society of Cell Biology. Kobe, Japan, 28–31 May 2012.
- Mochizuki, T., Suzuki, S., Sakaue-Sawano, A., Miyawaki, A. and Masai, I. Profile of cell cycle, cell division, and cell shape in lens: possible determinants of lens morphology. Joint meeting of the 45th annual meeting of the Japanese society of Developmental Biologists and the 64th annual meeting of the Japanese society of Cell Biology. Kobe, Japan, 28–31 May 2012.
Poster, domestic conferences
- Mochizuki, T., Suzuki, S., Sakaue-Sawano, A., Miyawaki, A. and Masai, I. E-cadherin controls apical cell geometry and orientation of cell division in the zebrafish lens epithelium. The 35th annual meeting of the Molecular Biology Society of Japan. Fukuoka, Japan. 11–14 December 2012.
- Yuasa-Kawada, J., Kinoshita-Kawada, M., Yanagi, S., Masai, I., Rao, Y., and Wu, J.Y. A switch required for axon crossing at the midline: crucial roles of Robo receptor endocytic trafficking and the deubiquitinase USP33. The 35th annual meeting of the Molecular Biology Society of Japan. Fukuoka, Japan. 11–14 December 2012.
- Kawada, J., Kinoshita-Kawada, M., Yanagi, S., Masai, I., Rao, Y., and Wu, J. Y. Upregulation of sensitivity to repellent Slit during midline crossing by commissural axons. Joint meeting of the 45th annual meeting of the Japanese society of Developmental Biologists and the 64th annual meeting of the Japanese society of Cell Biology. Kobe, Japan, 28–31 May 2012.
5. Intellectual Property Rights and Other Specific Achievements
Nothing to report
6. Meetings and Events
6.1 Developmental neurobiology course 2012
- Date: July, 16-30, 2012
- Venue: OIST Seaside House
- Co-organizers:
- Mary Ann Price (OIST)
- David L. van Vactor (Harvard Medical School, OIST)
- Robert Baughman (OIST)
- Akinao Nose (University of Tokyo)
- Speakers:
- David L. van Vactor (Harvard Medical School, OIST)
- Mary Ann Price (OIST)
- Susan McConnell (Stanford University)
- Mary E. Hatten (Rockefeller University)
- Ichiro Masai (OIST)
- Carol A. Mason (Columbia University)
- Alex Kolodkin (Johns Hopkins University)
- Jeniffer Waters (Harvard Medical School)
- Junying Yuan (Havard Medical School)
- Stephen Smith (Stanford University)
- Adrian W. Moore (RIKEN BSI)
- Matthew Dalva (Thomas Jefferson University)
- Anne Church Hart (Brown Research)
- Lesile Claire Griffith (Brandeis University)
- Yoko Sugiyama (OIST)
- Ryohei Yasuda (Max Planck Florida Institute)
- Akinao Nose (University of Tokyo)
- Arturo Alvarez-Buylla (University California San Francisco)
- Pierre Vanderhaegen (University of Brussels)